二萜合酶(DiTPS)是二萜生物合成分支中的第一个关键酶,催化二萜前体GGPP环化生成二萜骨架。被子植物通常利用II类和I类两个不同的单功能diTPS通过串联催化生成二萜骨架,首先由II类diTPS环化GGPP生成二环中间体,随后I类diTPS负责脱焦磷酸同时进行二次环化或骨架重排。然而,在二环二萜的生物合成中,是否由I类diTPS完成中间产物的脱磷酸步骤,一直存在争议。
半枝莲(Scutellaria barbata D. Don)为唇形科黄芩属多年生药用植物,富含克罗烷型二萜生物碱,目前已报道的有100多种,结构上主要由二环二萜模块和含氮模块通过酰基相连而成。这些化合物对多种肿瘤细胞表现出显著的细胞毒性,具有很大潜力成为抗肿瘤药物,而其生物合成途径未被解析。
2023年7月3日,澳门新葡萄新京威尼斯987黄莉莉课题组在国际学术期刊The Plant Journal在线发表了题为“Tandem duplication and sub-functionalization of clerodane diterpene synthase originates the blooming of clerodane diterpenoids in Scutellaria barbata”的研究论文。该工作组装了一个高质量的半枝莲基因组,在此基础上鉴定了半枝莲中参与克罗烷型二环二萜生物合成的两个II类二萜合酶基因,并提出I类二萜合酶不参与半枝莲中克罗烷型二萜生物合成的观点,该研究为克罗烷型二萜生物碱下游代谢途径解析和代谢工程研究奠定了良好的基础。

本研究采用以三代测序为主、二代测序辅助纠错、Hi-C挂载的基因组组装策略,组装了一个高质量的半枝莲基因组。基因组大小为322 Mb,98.8%的序列被挂载到13条染色体上(2n=26),Contig N50为18.40 M,Scaffold N50为29.42 M,共注释得到23,702个蛋白编码基因,BUSCO评估为97.3%。
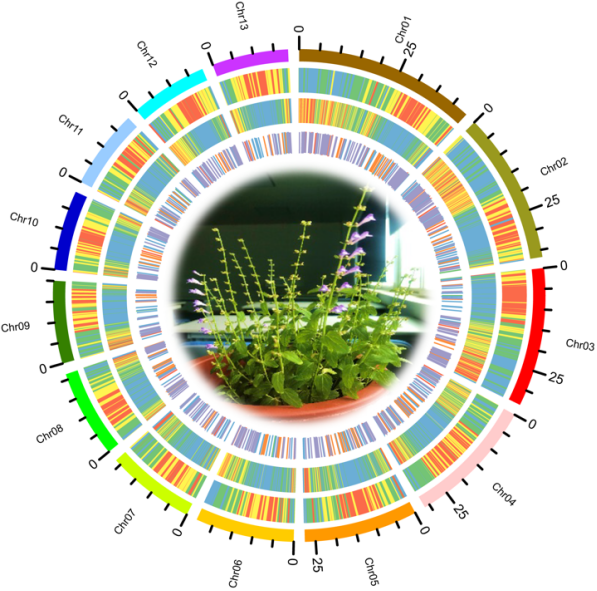
图1 半枝莲染色体水平基因组图谱
以基因组为基础,鉴定到两个克罗烷型II类diTPS SbarKPS1和SbarKPS2,蛋白序列同源性为91%,且在2号染色体上形成串联基因对。利用体外酶促、酵母和烟草异源系统证实SbarKPS1为单功能的克拉文基焦磷酸合酶((-)-KPS),SbarKPS2能同时生成neo-cleroda-4(18),13E-dienyl diphosphate和少量克拉文基焦磷酸。进一步通过比较基因组学发现,在同属药用植物黄芩中,仅有一个克罗烷型II类二萜合酶SbaiKPS1,生化功能与SbarKPS2一致。基于以上数据,我们推测串联重复和亚功能化导致了半枝莲中单功能克拉文基焦磷酸合酶SbarKPS1的产生,其产物进一步通过下游修饰酶的作用生成了半枝莲中结构多样的克罗烷型二萜生物碱。而黄芩中目前仅有一个克罗烷型二萜生物碱被报道。
在II类diTPS的基础上,进一步对下游的I类diTPS进行了鉴定,重点研究了功能未知的SbarKSL3和SbarKSL4。将其分别与不同的II类diTPS进行体外偶联酶促反应,结果表明,仅在不添加磷酸酶抑制剂的反应中,能检测到少量的脱磷酸产物,而添加了磷酸酶抑制剂之后,这些产物完全消失或者大幅下降,我们推测这些产物很可能来源于蛋白纯化过程中少量的大肠杆菌磷酸酶污染,而不是I类diTPS的产物。同时,在GGPP 高产的酵母底盘中过表达不同组合的II类diTPS和SbarKSL3/SbarKSL4,均未提高脱磷酸产物的产量。此外,qPCR结果表明SbarKSL3和SbarKSL4在各个组织中的表达量都很低,尤其是在叶中基本不表达,与上游II类diTPS SbarKPS1和SbarKPS2的表达模式及下游代谢产物半枝莲碱A和碱B的积累模式均不一致。基于以上研究结果,我们认为I类diTPS不参与半枝莲中克罗烷型二环二萜的生物合成,是否存在特异性催化这一步骤的酶有待于进一步研究。
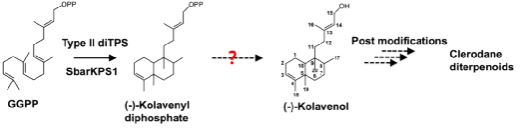
图2 半枝莲中克罗烷型二萜骨架克拉文醇(-)-Kolavenol生物合成途径示意图
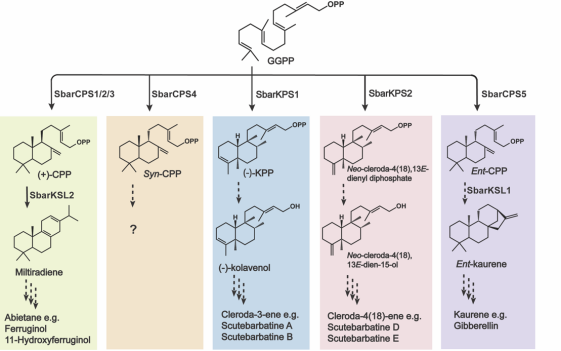
图3 半枝莲二萜合酶基因家族
该研究由澳门新葡萄新京威尼斯987药用植物生理生态研究所主导完成,澳门新葡萄新京威尼斯9872019级硕士研究生邱婷(已毕业)、2021级硕士研究生李洋演和吴海生为该论文的共同第一作者。澳门新葡萄新京威尼斯987黄莉莉研究员,申妍婷研究员和王宏斌教授为该论文的共同通讯作者。澳门新葡萄新京威尼斯987硕士研究生杨慧、彭梓萩、杜祖盈和博士研究生吴清文参与了相关研究工作。该研究得到了国家自然科学基金(32170400),国家自然科学基金区域创新发展联合基金(U22A20446)和广东省自然科学基金(2023A1515010895)等项目的资助。



 当前位置:
当前位置:







